Faktory klimatu / Kurz / Entomologie
Mezi faktory, které tvoří klima Země a jejích jednotlivých území, patří mezi hlavní z hlediska vlivu na živou přírodu sluneční záření, teplota a vlhkost vzduchu, k vedlejším pak vliv větru, ionizace a atmosférického tlaku a elektrických polí.
Solární radiace
Všechny procesy v biosféře zajišťuje sluneční záření, sluneční energie je však po zemském povrchu rozložena nerovnoměrně a navíc jsou pozorovány její sezónní a denní výkyvy, zvláště kontrastní v mírných a vysokých zeměpisných šířkách.
Sluneční záření určováním teploty vzduchu a různých substrátů vede ke změnám vlhkosti a atmosférického tlaku. Projevy téměř všech klimatických faktorů závisí na oslunění a následně mění intenzitu toku slunečního záření nad povrchem půdy.
Působení světla na hmyz se projevuje jednak fotosyntézou, při které vzniká organická hmota, jednak změnami dalších faktorů prostředí. Existuje však poměrně málo příkladů přímého vlivu světla na životně důležitou činnost hmyzu. Takže, larvy vážek Aeschna umbrosa L. ve tmě snižují rychlost metabolismu a po chvíli umírají, a housenky Antheraea pernyi G.-M. reagovat na změny světelných podmínek změnou aktivity některých enzymů. Význam světelného faktoru pro druhy aktivní pouze v noci a obyvatele jeskyní je omezený. Mezitím světlo, které je jasné pro lidské oko, může být pro hmyz slabé, ale konstantní jasné osvětlení ovlivňuje jeho životaschopnost, zejména životnost imaga. Existuje více příkladů vlivu světla na chování a vývoj hmyzu.
Hmyz může existovat více či méně dlouho v naprosté tmě. Světlo je však základem zrakových vjemů a různých biologických rytmů: denního (cirkadiánního), sezónního a lunárního. Intenzivní ultrafialové světlo je pro hmyz smrtelné.
teplota
Teplota je primární periodický faktor, který působí na živý organismus přímo a prostřednictvím změn jiných faktorů prostředí.
Vlastnosti povrchu předmětu – jeho barva, struktura, plocha – určují intenzitu absorpce a vyzařování tepla a hmotnost předmětů – jejich tepelnou kapacitu. Jak se velikost předmětu zmenšuje, jeho tepelná kapacita se mění rychleji než jeho schopnost absorbovat a přenášet teplo. V tepelném deficitu drsných podmínek vysokých zeměpisných šířek tak mají větší organismy oproti menším určité výhody.
Protože je hmyz velmi malý, na slunečních paprscích se ohřeje rychleji než ostatní živočichové, ale také rychleji vychladne ve stínu. Nemají stálou tělesnou teplotu, jsou mnohem více závislé na stavu prostředí a tepelné záření je pro ně významnější než teplota vzduchu.
Teplotní rozsah, ve kterém je možná aktivita hmyzu, se u různých druhů liší. Graf na obrázku 180 ukazuje, že toto rozmezí je omezeno na 15 a 38 °C a za tímto rozmezím intenzita životních procesů přirozeně klesá.
Teplotní optimum v zóně, kde jsou obecné projevy životní aktivity nejúčinnější s nejmenším výdejem energie, se posouvá směrem k vyšším teplotám. Suboptimální a superoptimální teplotní zóny umožňují normální aktivitu hmyzu, ale s dalšími změnami teploty nastává chlad nebo teplo, což vede k smrti. Když se tepelná necitlivost stane trvalou, návrat do aktivního života již není možný, ale ztuhlost z chladu je nahrazena podchlazením až do kritického bodu, kdy se uvolní latentní teplo tání a tělo hmyzu se zahřeje téměř na 0 °C. Některé druhy hmyzu jsou schopny vydržet velmi dlouhé a hluboké poklesy teploty ve stavu pozastavené animace. Například housenky zavíječe kukuřičného (Ostrinia nubilalis L.) ožil po ochlazení na -190 °C. Při vysvětlení tohoto jevu a výpočtu kritického bodu podchlazení je třeba vzít v úvahu, že šťávy z těla hmyzu slouží jako nemrznoucí směs a zamrzají při teplotách nižších než voda.
V aplikovaném výzkumu se často setkáváme s potřebou stanovit meze tolerance a teploty preferované hmyzem. Dospělí můry cikánské (Ocneria dispar L.), zemřou například při půlhodinovém ochlazení na -4 °C. Imago kobylky Melanoplus mexicanus L. hynou při -8 °C po dvou dnech a zimující housenky zlatoocasého: Nygmia phaeorrhoea L. jsou schopny vydržet ochlazení na -14 °C po dobu 158 dní.
Když teplota stoupne na 45–48 °C, mnoho hmyzu uhyne, ale larvy některých drahokamů a nymf pouštních sarančat zůstávají aktivní i při 52 °C a larvy pobřežních mušek jsou aktivní i při 65 °C.
Teploty běžné pro mnoho druhů hmyzu mají k těmto extrémům daleko. Schopnost hmyzu snášet zvýšené nebo snížené teploty se liší v závislosti na fyziologickém stavu, předchozích zkušenostech a životních podmínkách jedinců.
Při identifikaci preferovaných teplot (termopreferendum) se obvykle používají zařízení, která udržují tepelný gradient nebo poskytují možnost vybrat jednu ze dvou experimentálních komor vyhřívaných na různé stupně. Za těchto podmínek moucha domácí (Musca domestica L.) obvykle preferuje teplotu 42 °C, veš Pediculus humanis L. — od 24 do 32 °C, ale hodnoty tepelné preference se mohou měnit v závislosti na doprovodných okolnostech. Například mravenci Formica rufa L., dříve udržované na 3–5 °C, volily teploty od 23 do 29 °C a ty, které byly udržovány na 27–29 °C, volily teploty od 32 do 52 °C. Pro kobylku pouštní (Schistocerca gregaria Forsk.) byl zjištěn následující průběh změn tepelné preference, °C: nymfa prvního věku – 30,1, nymfa druhého věku – 28,8, nymfa třetího věku – 31,6, nymfa čtvrtého věku – 37,1, nymfa pátého věku – 36,7, mladí dospělí – 39,3, pohlavně zralí – 29,4. V noci mnoho hmyzu preferuje nižší teploty než přes den.
Podle teplotní preference, která vyjadřuje biologické nároky druhu, a rozmezí tolerovaných teplot se hmyz dělí na stenotermní a eurytermní. První mohou tolerovat pouze omezené výkyvy teplotních podmínek, zatímco druhé jsou schopny odolávat změnám faktoru v širších mezích. Paraziti teplokrevných živočichů, stejně jako mnozí zástupci tropické fauny, jsou obvykle stenotermní a teplomilní. Mezi stenothermní chladnomilné druhy himálajské sněžné fauny patří skupiny ocásků a dvoukřídlých, kteří jsou aktivní v teplotním rozmezí 0 až 10 °C, 0 až 5 °C a nakonec aktivní pouze během slunečných hodin dne při teplotách 5 až 10 °C. Zástupci prvních dvou skupin jsou tak citliví na zvýšení teploty, že zemřou během několika minut z tepla lidské ruky.
Příklady eurytermálních druhů: mucholapka zelená (Lucilia serricata Mg.), motýl želvovinový (Aglais urticae L.), obrněnci (Oribatei), žijící na holých skalách ve vnitrozemí a tolerující výkyvy od 60 °C ve dne do 0 °C v noci.
Hmyz má také svoji termoregulaci. Svědčí o tom například fakt, že mrtví potemníci v poušti se zahřívají o 2–9 °C více než živí. Černý šváb (Blatta orientalis L.) v důsledku odpařování z povrchu těla nebo zvýšené spotřeby kyslíku se teplota buď mírně zvýší, nebo sníží, ale pouze v případě, že okolní teplota překročí 13–25 °C.
V horkém počasí včely větrají úl, ale při teplotách pod 13 °C se začnou energicky pohybovat a choulit se k sobě. Zároveň se jedinci ochlazující se na periferii shluku plazí do jeho středu a vytlačují ty, kteří se oteplili. V důsledku toho teplota vzduchu v úlu stoupne na 30 °C. Velký význam pro termoregulaci má chvění křídel čmeláků a jestřábů, kteří jsou schopni vzlétnout, až když teplota křídelních svalů není nižší než 34,5 °C. Mnoho druhů hmyzu zvyšuje svou teplotu tím, že se plazí ven do osluněných oblastí.
Pro posouzení vlivu teploty na průběh vývoje hmyzu se obvykle používá kritérium součtu efektivních teplot, jehož počítání se provádí z dříve vypočtených hodnot spodního prahu vývoje. Například ovocná muška (Drosophila melanogaster Mg.) se vyvíjí při teplotách ne nižších než 13,5 °C, což je prahová hodnota pro tento druh. Odpovídající prahové hodnoty teploty pro mšice jablečné (Aphispomi Deg.), moucha domácí (Musca domestica L.) a červený šváb (Blatella germanica L.) jsou 7,12 a 6,5 °C. Pro sýpku (Sitophilus granaria L.) teplotní práh pro vývoj je 11 °C, u molice zelné (Plutella maculipennus Curt.) – 9,8, švédská muška (Oscinella frita L.) – 8, zimní můra (Agrotis segetum Schiff.) – 10, mandelinka bramborová (Leptinotarsa decemlineata Say.) — 12 °C.
Při nižších teplotách se vývoj zpomaluje, ale pokud jejich průměrné denní hodnoty překročí práh, pak vývoj pokračuje a hmyz dosáhne pohlavní dospělosti po požadované době.
V mezích přijatelných pro každý druh se s rostoucí teplotou zkracuje doba vývoje a zvyšuje se jeho rychlost. Tato závislost, vyjádřená Blunckovou hyperbolou, odpovídá principu kinetiky chemických reakcí, podle kterého zvýšení teploty o 10 °C vede k 2–4násobnému zrychlení reakcí a procesů. Embryonální vývoj mandelinky bramborové při 17 °C je tedy dokončen po 216 hodinách a při 27 °C po 96 hodinách.
Schopnost biologické. objektů termoregulace, aktivního výběru preferovaných teplot a udržování homeostázy nám umožňuje porovnat termostabilní a termolabilní procesy, fáze a fáze, které jsou v menší či větší míře náchylné na vliv teploty. Zejména období štěpení v embryogenezi mandelinky bramborové je relativně termostabilní a jeho konečná stádia jsou termolabilní.
Schopnost brouka vojáka, který je v Evropě rozšířený,Pyrrhocoris apterus L.) produkovat pouze jednu generaci jak na jihu, tak na severu rozsahu je určeno selektivitou substrátů zahřátých na maximálně 23 °C.
Výpočet součtů efektivních teplot (S) se provádí podle vzorce:
kde t — průměrná denní teplota vzduchu; t — hodnota spodního prahu rozvoje; n — počet dní potřebných pro vývoj.
Například, Drosophila melanogaster Mg, (t = 13,5 °C) při 26 °C se vyvine za 20 dní a při 19,5 °C – 41,7 dní. Tedy v prvním případě, kdy S= (26 – 13,5)20 = 250, a ve druhém – S = (19,5 – 13,5) * 41,7 = 250,2 součet efektivních teplot je stejný (tepelná konstanta) a lze jej použít k predikci doby vývoje, od r. n = S/(t – t).
U mšic jablečných součet efektivních teplot (S), °C — 114, pro mouchu domácí — 230, pro sýpku — 360, pro můru zelnou — 380, pro můru fritovou — 400, pro můru zimní — 1 000, pro švába rudého — 1900. Například s vědomím, že součet teploty v oblasti zelí je přibližně 9°C nad 740 °C °C, zde lze očekávat výskyt minimálně dvou generací škůdce. V Omské oblasti (S = 1 150 °C) je možný vývoj tří generací tohoto škůdce v oblasti Krasnodar (S = 2 300 °C) — již 5–6 generací. Takové výpočty jsou použitelné pouze pro ty druhy, jejichž počet generací se liší v závislosti na podmínkách existence a není geneticky fixován, jako například u bource morušového. Bombyx mori L.
Zaznamenávání součtů efektivních teplot nám často umožňuje předvídat načasování výskytu škodlivých fází a plánovat ochranná opatření. Zejména je vhodné použít insekticidy proti mandelince bramborové na bramborách, když S = 150 °C (at t = 12 °C), protože v této době začíná hromadný výskyt škůdce.
Výpočty součtů efektivních teplot mají přibližnou hodnotu; Měly by být upraveny s ohledem na údaje z přímých pozorování stavu populací škůdců. Doba vývoje škůdců navíc závisí nejen na akumulovaném teple, ale také na výkyvech teplot, které většinou přispívají k jejich snížení. Při nižších teplotách se výrazně prodlužuje životnost jednotlivých jedinců. Tedy pokud při 30 °C letí Drosophila melanogaster Mg. existovat v průměru 21 dní, poté při 15 °C – minimálně 130 dní.
Plodnost hmyzu ovlivňují i klimatické faktory: je maximální za optimálních podmínek, ale celoživotní plodnost samic závisí na délce jejich existence. Mnohdy je snížení počtu vajíček denně snesených samicí při nižších teplotách kompenzováno délkou snáškového období a v konečném důsledku jsou nejplodnější jedinci nejdéle žijící, nikoli nejaktivnější. Nejvíce vajíček za jednotku času nakladou samičky mouchy švédské při teplotě 18–39 °C, moucha meromyza (Megomyza saltatrix L.) – ve 20-30 a můra luční Loxostege sticticalis L. – při 25 °C.
Влажность
Je obtížné a někdy nemožné oddělit vliv vlhkosti od vlivu jiných faktorů prostředí. Pokud jsou k dispozici zdroje vlhkosti, hmyz snadno snáší suchý vzduch a na souši umírají pouze formy, které neustále žijí ve vodních útvarech (hydrobionti).
Vliv vlhkosti na dobu trvání a rychlost vývoje je obvykle určen vlivem teploty. U much domácích a zelených muchniček (Musca domestica L. a Lucilia serricata Mg.) byl zjištěn lineární vztah mezi rychlostí vývoje a vláhovým deficitem a u sarančat stěhovavých se rychlost vývoje zvyšuje se zvýšením relativní vlhkosti na 10 %. Při vyšší vlhkosti je vývoj sarančat brzděn a mnoho jedinců je postiženo houbovými a bakteriálními chorobami.
Suchozemský hmyz se dělí na hygrofily, kteří žijí na vlhkých místech, mezofily méně náročné na vzdušnou vlhkost a xerofily, kteří jsou adaptováni na neustálý nedostatek vlhkosti.
Aby si doplnili zásoby vody, pije některý hmyz rosu (želva ploštice – Eurygaster integriceps Pot.) nebo rostlinné šťávy, konzumovat vlhkost s jídlem nebo využívat metabolickou vodu vznikající při oxidaci tuků a jiných organických látek. Housenky mola šatního (Tineola biselliella L.) a můra mlýnská (Ephestia kuhniella Zell.), sýpka a rýžové nosatce (Sitophilus granaria L. a S. oryzae L.), krmení na substrátech s nízkým obsahem vlhkosti, vystačí s metabolickou vodou. Mezitím hygrofilní hmyz a zejména vodní organismy dýchající žábry spotřebovávají značné množství vody prostřednictvím propustných vrstev.
Jednou z možností, jak ušetřit zásoby vody, které jsou neustále spotřebovávány při metabolismu, transpiraci a vypařování, je vylučování koncentrované moči. Filtrační komory střev slouží k odstranění přebytečné vody spotřebované rostlinnou šťávou.
Různé druhy hmyzu snášejí snížení obsahu vody v těle různě: mandelinka bramborová (Leptinotarsa decemlineata Say.) odolává vysychání o 50 %, brouk Catalpa lanigera L. — o 25 %, a brouk Lachnosterná sp. po vysušení zemře o 15 %. Mezitím larvy některých komárů, žijící v vysychajících kalužích, zůstávají naživu 10 let a ve vodě si neudrží více než 3 % své tělesné hmotnosti. Tyto údaje však charakterizují rozmanitost reakcí hmyzu na změny vlhkosti pouze za experimentálních podmínek.
V přírodě je vliv vlhkosti zprostředkován teplotou a dalšími faktory prostředí. Proto se v praxi environmentálního výzkumu obvykle uvažuje o kombinovaném vlivu vlhkosti a teploty, k jehož demonstraci se používají různé indexy, koeficienty a diagramy. Při jejich výpočtu a konstrukci je vhodné použít parametry akceptované v meteorologii: průměrné (denní průměr, průměr za desetiletí atd.) hodnoty teploty a vlhkosti vzduchu s vlhkostí zohledněnou buď v relativním vyjádření, nebo jako množství srážek za dané období.
K predikci a identifikaci limitujících faktorů pro každou fázi vývoje hmyzu se používají bioklimogramy B. P. Uvarova (obr. 181).
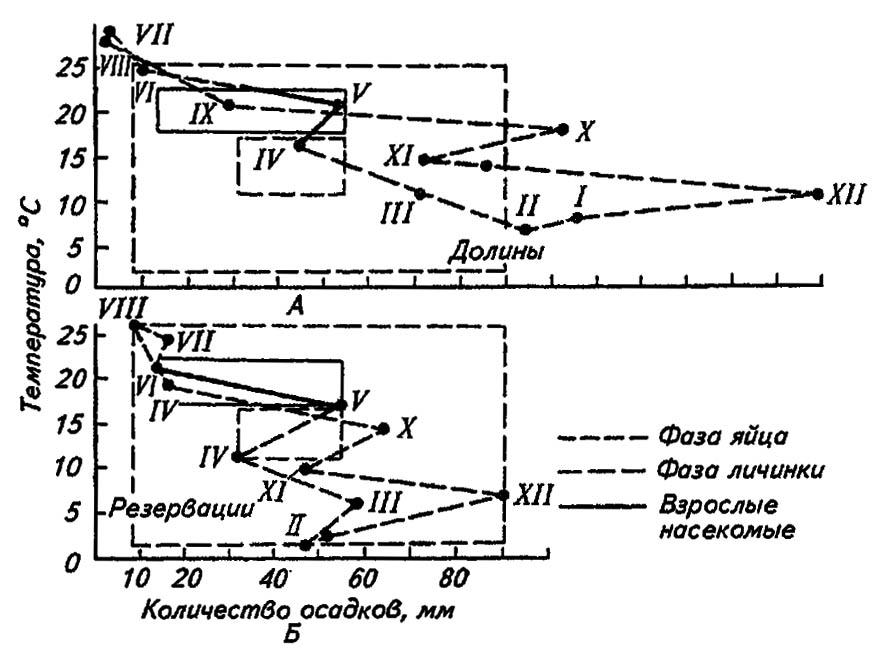
Obrázek 181. Bioklimogram B. P. Uvarova pro kobylku marockou v Malé Asii (podle Bey-Bienko, 1966):
А – nepříznivé a Б – příznivé životní podmínky
S meteorologickými zprávami pro území a období hromadného výskytu druhu je možné na odpovídajícím grafu načrtnout optimální zónu, jejíž obrysy při superponování do grafů vytvořených pro jiná území a období okamžitě odhalí, co a pro jakou fázi vývoje slouží jako omezující faktor. Z poskytnutého grafu je zřejmé, že příčinou úhynu zimujících vajíček saranče marockého byly nadměrné srážky v říjnu (X), prosinci (XII), lednu (I) a únoru (II), stejně jako jejich nedostatek a zvýšení teploty v létě (VI–VIII). Pokles teplot v dubnu (IV.) měl nepříznivý vliv na dospělé.
Vedlejší faktory
Při silném větru se hmyz, který se nestihne uchýlit do úkrytů, přenáší na velké vzdálenosti. Když se ocitnou v neznámých podmínkách, umírají nebo se těmto podmínkám přizpůsobují a eurytopní druhy, které se vyznačují zvýšenou ekologickou valencí, mají větší šanci na přežití než stenotopní druhy. Podmínky v nové domovině jsou pro migranty jen zřídka příznivé a hmyz se většinou vyhýbá poryvům větru. Některé z nich, které mají zvláštní plachty – načechrané štětiny a výrůstky těla, jsou však přizpůsobeny pasivnímu rozptýlení vzdušnými proudy.
Při prudkém poklesu atmosférického tlaku, který obvykle věští bouři, je aktivní zejména hmyz. To lze snadno ověřit pozorováním mraveniště. Později se přestanou pohybovat a sníží svou aktivitu, čekají na hurikán, ale zůstávají vystaveni nepřímým vlivům větru, který mění teplotu a podporuje odpařování vlhkosti. Snížený atmosférický tlak stimuluje výskyt některých druhů motýlů a potápníků. Kromě toho se zdá, že hmyz při migraci sleduje pokles tlaku a spěchá do oblastí, kde jsou jeho hodnoty nižší. Jsou také náchylné k atmosférické ionizaci před a po bouřkách, při lesních požárech a elektrických výbojích blesku.
Pozitivní ionty zpravidla stimulují aktivitu hmyzu, jsou lhostejné k negativním, ale v laboratorních experimentech mšice Acyrthosiphon pisi Kalt. reagoval na ně inhibicí línání. Mnoho hmyzu je ovlivněno změnami směru elektrického pole, pravděpodobně v důsledku akumulace elektrostatických nábojů na těle. Při geomagnetických bouřích se zvyšuje aktivita mnoha hmyzu a ovocné mušky reagují i na slabé elektrické pole (do 200 V/m) a jeho změny, například když je přelétají mraky.